
 Light does not reach the deep-sea floor. This precludes photosynthesis and thus primary production (except in chemosynthetic communities like hydrothermal vents and methane seeps). The lack of primary production on the deep-sea floor results in these communities being intimately coupled to the production of food at the surface by phytoplankton. What fluxes down to the deep sea is a combination of decaying animals (from plankton to whales), fecal pellets, and photosynthetic material (phytoplankton to seaweed). This material (carbon compounds) is consumed by deep-sea species and either returned to the water column as carbon dioxide from respiration or enters into the sediment. This relationship between the seafloor and surface is termed benthic-pelagic coupling with the actual process of carbon movement from the surface to the floor called carbon flux. Carbon flux and benthic-pelagic coupling both fit into the broader concept of carbon cycling with terrestrial plant production and the consumption of fossil fuels.
Light does not reach the deep-sea floor. This precludes photosynthesis and thus primary production (except in chemosynthetic communities like hydrothermal vents and methane seeps). The lack of primary production on the deep-sea floor results in these communities being intimately coupled to the production of food at the surface by phytoplankton. What fluxes down to the deep sea is a combination of decaying animals (from plankton to whales), fecal pellets, and photosynthetic material (phytoplankton to seaweed). This material (carbon compounds) is consumed by deep-sea species and either returned to the water column as carbon dioxide from respiration or enters into the sediment. This relationship between the seafloor and surface is termed benthic-pelagic coupling with the actual process of carbon movement from the surface to the floor called carbon flux. Carbon flux and benthic-pelagic coupling both fit into the broader concept of carbon cycling with terrestrial plant production and the consumption of fossil fuels.
Most benthic-pelagic coupling models focus primarily on the dynamics of carbon cycling at the ocean surface not at the seafloor. These models incorporate dynamics of phytoplankton growth, zooplankton grazing, and microbial utilization in surface waters to assess flux into and out of photic zone. How much carbon is fluxed is dictated by dynamics between phytoplankton and zooplankton. If zooplankton consume all the phytoplankton little carbon will reach the deep sea. However, when phytoplankton bloom there is a lag in time for the zooplankton respond. During the time it takes for the zooplankton population to increase in numbers to consume all the phytoplankton, the extra material sinks to the ocean bottom.
Overall, approximately 25% (16 Gt/yr) of carbon fixed by phytoplankton is thought to sink to the ocean interior (Falkowski 1998). Compare this 16,000,000,000 tons to the total carbon dioxide emissions, 25,167,070 tons, from consumption of fossil fuels for 2003. This exemplifies the importance of a better understanding of how deep-sea benthic communities contribute to benthic-pelagic coupling in elucidating the dynamics of the carbon cycle.
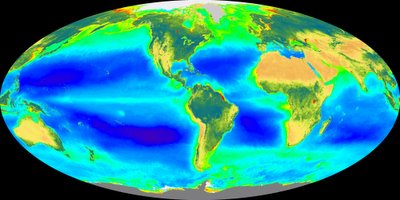
As one would expect patterns and processes in deep-sea communities are largely controlled by the magnitude and variability of organic carbon flux from the euphotic zone. To some extent the magnitude of flux mirrors the amount of total production at the seasurface which typically decreases with distance from land. The map above shows where the highest ocean production (warm colors). The amount of carbon arriving at the seafloor also depends on the the depth of the water column. The longer the material is in the pelagic zone (increasing depth) the more likely it is to be consumed by pelagic species or degraded by bacteria in the water column. Several studies have shown a variety of features to be linked to this flux including:
- geographic gradients in species diversity (Rex 1973, Rex et al. 1997)
- standing biomass (Rowe 1983)
- abundance (Rex 1989)
- body size (Thiel 1975, McClain et al. 2005)
- migration (Bett et al. 2001)
- reproduction (Gage and Tyler 1991)
- metabolism (Rowe 1996).
Studies also demonstrate that deep-sea communities can respond to pulsed fluxes of primary production from the euphotic zone at time scales of less than 2 weeks (Graf 1989, Gooday et al. 1990, Smith et al. 2002) but over geological time periods as well (Cronin and Raymo 1997)
Share the post "ARCHIVE: 25 Things You Should Know About the Deep Sea: #8 Processes & Patterns In the Deep-Sea Are Linked to Surface Production"



The lack of primary production on the deep-sea floor results in these communities being intimately coupled to the production of food at the surface by phytoplankton.
Does “lack” imply nonexistence of primary production on the deep sea floor? Cause we know that’s not the case — chemoautotrophs are necessary for many deep sea environments.
Also, the last section of your post, following “including:” is not rendering correctly in my browser (Firefox 1.5). The text is there, but I can only read it if I highlight it with my cursor.
For most of the deep sea, probably greater than 95%, there is no primary production. As you note there are exceptions, hydrothermal vents and methane seeps, where chemautotrophy drives an ecosystem. As pervasive as they are there impact on total food input for the deep sea is quite low. Thanks for the heads up on the wording will fix.